Кому принадлежит описание фаз мейоза. Определение и виды мейоза
Николай Мушкамбаров, докт. биол. наук
Человечество стареет, а ведь каждому хочется жить не просто долго, но и без тех болезней, которые приходят с возрастом. За последние полвека появилось множество «революционных» теорий старения, почти каждая из которых предлагает верный и надёжный способ замедлить или даже остановить время. Каждый год - новые сенсации, новые открытия и новые заявления, обнадёживающие и многообещающие. Пептидные биорегуляторы, эликсир долголетия, живительные ионы, или антиоксидант SkQ. Беги в аптеку, плати и живи, согласно вложенной инструкции, до 100-120 лет! В какой мере можно доверять сенсационным открытиям и в чём состоит «правда о старении»?
Профессор Н. Н. Мушкамбаров. Фото Андрея Афанасьева.
Август Вейсман (1834-1914) - немецкий зоолог и эволюционист. Создал теорию, согласно которой наследственные признаки сохраняются и передаются через нестареющую зародышевую плазму.
Леонард Хейфлик - американский микробиолог. В 1960-е годы обнаружил, что в лабораторных условиях клетки человека и животных могут делиться лишь ограниченное число раз.
Алексей Матвеевич Оловников - российский биохимик. Для объяснения экспериментов Хейфлика в 1971 году выдвинул гипотезу об укорачивании концевых участков хромосом (теломер) при каждом делении клетки.
Наука и жизнь // Иллюстрации
Элизабет Блэкбёрн и Кэрол Грейдер - американские биологи. В 1985 году открыли фермент теломеразу. Механизм действия теломеразы - повторяющееся кодирование новых нуклеотидных последовательностей на концевых участках теломер и восстановление их исходной дл
Бенджамин Гомперц (1779-1865) - британский математик. Предложил функцию, описывающую статистику смертности человека в зависимости от возраста. Эта функция использовалась для оценки рисков при страховании жизни.
Книга М. М. Виленчика «Биологические основы старения и долголетия», изданная в 1976 году, была одной из первых научно-популярных книг на тему старения и пользовалась огромным успехом.
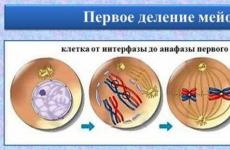
Схема мейоза (на примере пары гомологичных хромосом). В профазе первого деления мейоза хромосомы удваиваются; затем гомологичные хромосомы конъюгируют друг с другом и, сохраняя свою активность, вступают в кроссинговер.
На вопросы специального корреспондента журнала «Наука и жизнь» Наталии Лесковой отвечает доктор биологических наук, профессор кафедры гистологии Московского государственного медицинского университета им. И. М. Сеченова Николай Мушкамбаров.
Николай Николаевич, вы резко критикуете многие получившие широкую известность положения современной геронтологии. Пожалуйста, обрисуйте объекты вашей критики.
Объектов хоть отбавляй! Например, сейчас модно ссылаться на Вейсмана чуть ли не как на истину в последней инстанции. Это знаменитый биолог, который ещё в XIX веке постулировал, что старение возникло в эволюции не сразу, а лишь на каком-то её этапе как приспособительное явление. Отсюда сделали вывод, что должны быть нестареющие виды: в первую очередь наиболее примитивные организмы. При этом как-то забывают, что если они не стареют, то у них должна быть 100%-ная репарация ДНК. Это у самых-то примитивных! Как-то не вяжется одно с другим.
Есть миф, связанный с именем другого известного биолога - Леонарда Хейфлика. С шестидесятых годов прошлого века научный мир пребывает в уверенности, что соматическим клеткам человека положен предел в 50 делений, и такой лимит в биологии называют «пределом Хейфлика». Лет двадцать назад выделили стволовые клетки, способные якобы к неограниченному числу делений. И этот миф (50 у всех и бесконечность у стволовых клеток) сохраняется в умах и поныне. На самом же деле и стволовые клетки, как выясняется, стареют (то есть бесконечность отменяется), и совсем непонятно, откуда вести отсчёт этих самых 50 делений. Настолько непонятно, что, скорее всего, какого-то одного лимита делений, универсального для всех делящихся клеток человека, не существует.
- Ну а теломерная теория старения? Она тоже вызывает у вас недоверие?
Это самый популярный миф. Согласно этой теории, весь механизм старения сводится к тому, что в делящихся клетках отсутствует фермент теломераза, удлиняющий концы хромосом (эти концы называются теломерами), и потому при каждом делении происходит укорочение теломер на 50-100 нуклеотидных пар ДНК. Фермент теломераза действительно существует, и его открытие отмечено Нобелевской премией 2009 года. И феномен укорочения хромосом в делящихся клетках, лишённых теломеразы, тоже не вызывает сомнения (хотя обусловлен немного не той причиной, на которую указывал автор теломерной теории Алексей Оловников). Но сводить к этому феномену старение - всё равно, что заменить сложнейшую партитуру симфонии нотами стука по барабану. Неслучайно в 2003 году А. Оловников публично от своей теории отказался, заменив на так называемую редумерную теорию (тоже, кстати, не бесспорную). Но до сих пор даже в медицинских вузах в курсе биологии излагают теломерную теорию в качестве последнего достижения научной мысли. Это, конечно, абсурд.
Ещё один пример - из статистики смертности. Главная формула этой статистики - уравнение Гомперца, предложенное в 1825 году, или, с поправочным членом, уравнение Гомперца-Мейкема (1860 г.). В этих уравнениях соответственно есть два и три коэффициента, причём значения коэффициентов сильно варьируют для разных популяций людей. И вот, оказывается, изменения коэффициентов каждого уравнения коррелируют друг с другом. На основании чего формулируются глобальные, общемировые закономерности: так называемая корреляция Стрелера-Милдвана и сменивший её на этом посту компенсационный эффект смертности - гипотеза супругов Гавриловых.
Я же составил небольшую модель для условной популяции людей и с её помощью убедился, что все эти закономерности, скорее всего, артефакт. Дело в том, что небольшая погрешность в определении одного коэффициента создаёт резкое отклонение от истинного значения другого коэффициента. А это воспринимается (в полулогарифмических координатах) как биологически значимая корреляция и служит посылом для глубокомысленных заключений.
- Уверены ли вы в своей правоте, говоря об артефакте?
Конечно, нет! Учёным вообще вредно быть в чём-то абсолютно уверенными, хотя таких примеров предостаточно. Но я сделал всё, что в силах, чтобы проверить обратное: то, что корреляции не артефакт. И вот убедиться в этом обратном мне не удалось. Так что пока на основании личного, очень скромного по масштабу, анализа у меня больше оснований считать, что названные корреляции всё-таки искусственные. Они отражают погрешности метода, а не биологические закономерности.
А как вы оцениваете заявления о том, что в природе существует огромное количество нестареющих организмов и их список год от года растёт?
Увы, популярные теории о том, что существуют как нестареющие клетки, так и нестареющие организмы, лишены достаточных оснований. Действительно, с каждым годом круг «нестареющих» животных неумолимо расширяется. Вначале это были практически только одноклеточные, затем к ним добавили низших многоклеточных (гидр, моллюсков, морских ежей и др.). А теперь появились горячие головы, которые «обнаруживают» отдельные нестареющие виды даже среди рыб, рептилий и птиц. Так пойдёт - скоро доберутся и до млекопитающих и установят, например, что и слоны тоже не стареют, а погибают просто из-за избыточной массы тела!
- Вы убеждены, что нестареющих животных нет?
Я убеждён не в том, что таких животных нет (хотя и склоняюсь к этому), а в том, что нет ни единого вида животных, для которых отсутствие старения было бы доказано абсолютно надёжно. В отношении же клеток человека (равно как клеток и прочих представителей животного мира) степень уверенности, пожалуй, ещё выше: и стволовые, и половые, и даже опухолевые клетки, в принципе, стареют. Уж куда как бесспорно нестареющими считались стволовые клетки, - а теперь появляются экспериментальные работы, доказывающие противное.
- На чём основывается такая уверенность? Вы сами проводили соответствующие эксперименты?
Вообще говоря, очень давно, в 1977-1980 годах, я пытался подступиться к проблеме старения в экспериментах на мышах. Но не очень надёжные результаты (хотя вроде бы и подтверждающие исходное предположение) убедили, что лучше всё-таки заниматься не экспериментом, а анализом. И вот один из итогов этого анализа - концепция «Анерем», или амейотическая теория старения. Она включает шесть тезисов (если хотите, постулатов), из которых один (самый первый) - чисто моё творчество, а остальные сформулированы на основании уже имевшихся в литературе представлений. Ну и, конечно, важно, что все эти тезисы составили в целом достаточно чёткую картину.
Так вот, именно амейотическая концепция, если её придерживаться, исключает возможность существования и нестареющих клеток в многоклеточных организмах, и нестареющих организмов (начиная с одноклеточных). При этом, конечно, я отдаю себе отчёт, что все тезисы концепции - пока ещё гипотезы. Но они представляются гораздо более обоснованными, чем прочие взгляды.
Значит, ваша концепция вроде тестера, с помощью которого можно оценить, условно говоря, истинность тех или иных предположений? В таком случае расскажите о ней подробнее.
Постараюсь сделать это максимально доступно. Само название концепции («Анерем») представляет собой аббревиатуру от слов автокатализ, нестабильность, репарация, мейоз. Тезис первый. Помните, прежде было очень известно определение жизни по Энгельсу: «Жизнь есть способ существования белковых тел»? Я пересмотрел это определение и дал своё, которое и составило первый тезис: «Жизнь есть способ автокаталитического умножения ДНК (реже РНК) в природе». Это значит, что движущая сила и возникновения жизни, и последующей её эволюции - это неукротимое стремление нуклеиновых кислот к бесконечному самовоспроизведению. По существу, любой организм - усовершенствованная в эволюции биомашина, предназначенная для эффективного сохранения и умножения содержащегося в нём генома с последующим эффективным распространением его копий в окружающей среде.
- Непривычно ощущать себя биомашиной…
Ничего, ощущение пройдёт, а функция, простите, останется. Тезис второй: «Нестабильность генома - центральный элемент старения». Именно так понимают старение большинство здравомыслящих учёных на Западе, да и у нас. Дело в том, что, при всех своих замечательных способностях, нуклеиновые кислоты подвержены повреждающему воздействию множества факторов - свободных радикалов, активных форм кислорода и т.д. И хотя в эволюции было создано немало защитных систем (таких, как антиоксидантная система), в нитях ДНК постоянно возникают многочисленные повреждения. Для их обнаружения и исправления существует ещё одна защитная система - репарации (восстановления) ДНК. Следующий тезис, третий, - это фильтр, который отсеивает всё «нестареющее»: «Репарация генома в митотических и постмитотических клетках не является полной». То есть любая система репарации в указанных клетках не обеспечивает 100%-ного исправления всех возникающих дефектов ДНК. А это и означает всеобщий характер старения.
- Но если всё и вся стареет, то как же вообще поддерживается жизнь на Земле?
Вот-вот, этим вопросом заинтересовался и я в 1977 году. И нашёл, как мне показалось, свой собственный, хотя и лежащий на поверхности, ответ. А через 25 лет, в 2002-м, перебирая свои старые книги, понял, что гипотеза эта вовсе не моя, а я о ней читал за год до того в книге М. М. Виленчика, благополучно забыл и потом вспомнил, но воспринял как свою собственную. Вот такие причуды памяти. Но, в конце концов, важна суть дела, а не амбиции первооткрывателя.
Суть же формулируется тезисом четвёртым: «Эффективная репарация может быть достигнута только в мейозе (или в его упрощённом варианте - эндомиксисе) - при конъюгации (слиянии) хромосом». Что такое мейоз, все вроде бы проходили в школе, но, к сожалению, этого не знают порой даже наши студенты-медики. Напоминаю: мейоз - это последнее двукратное деление при образовании половых клеток - сперматозоидов и яйцеклеток. Кстати, по секрету сообщу: у женщин яйцеклеток не образуется. У них второе мейотическое деление (на стадии ооцита II - развития женской половой клетки) не может происходить самостоятельно - без помощи сперматозоида. Потому что клетка куда-то «затеряла» свои центриоли (тельца в клетке, участвующие в делении): только что (при предыдущем делении) были, а теперь вот куда-то делись. И требуется непременно оплодотворение ооцита II, чтобы сперматозоид привнёс свои центриоли и спас положение. Я это рассматриваю как типичные «женские штучки». Так что второе деление мейоза в конце концов происходит, но образующаяся в результате клетка - это уже не яйцеклетка, а зигота.
Мы увлеклись «женскими штучками» и не прояснили, каким же образом достигается полная репарация ДНК в мейозе.
Первому делению мейоза предшествует очень длительная профаза: в мужском гаметогенезе она продолжается целый месяц, а в женском - до нескольких десятков лет! В это время гомологичные хромосомы сближаются друг с другом и в таком состоянии пребывают почти всё время профазы.
При этом резко активируются ферменты, разрезающие и сшивающие нити ДНК. Считалось, что это необходимо только для кроссинговера - обмена хромосом своими участками, что увеличивает генетическую вариабельность вида. Действительно, «папины» и «мамины» гены, до сих пор распределённые в каждой паре гомологичных (одинаковых структурно) хромосом по разным хромосомам, после кроссинговера оказываются перемешанными.
Но М. М. Виленчик, а вслед за ним и я, обратил внимание на то, что ферменты кроссинговера уж очень напоминают ферменты репарации ДНК, при которой, вырезая повреждённые участки, тоже надо разрывать и сшивать нити ДНК. То есть одновременно с кроссинговером, вероятно, осуществляется суперрепарация ДНК. Можно представить и другие механизмы капитального «ремонта» генов в ходе мейоза. Так или иначе, при этом происходит радикальное (точнее, полное) «омоложение» клеток, отчего зрелые половые клетки начинают отсчёт времени как бы с нуля. Если же что-то не получилось, то в клетке срабатывают датчики самоконтроля за состоянием собственной ДНК и запускается процесс апоптоза - само-
убийства клетки.
- Значит, в природе омоложение происходит только у созревающих половых клеток?
Совершенно верно. Но этого вполне достаточно для обеспечения бессмертия вида - на фоне, увы, неотвратимой смертности всех индивидов. Ведь половые клетки - и только они! - тот единственный материальный субстрат родительских организмов, из которого зарождается новая жизнь - жизнь потомства.
И о том, что этот механизм касается лишь половых клеток, идёт речь в двух оставшихся тезисах концепции, которые расставляют все точки над i. Тезис пятый: «Мейоз улучшает состояние генома только последующих поколений (сразу нескольких поколений у простых организмов и лишь одного у всех прочих)». Тезис шестой: «Отсюда следуют неотвратимость старения индивидуумов (особей) и относительное бессмертие вида в целом».
- А что, мейоз есть у всех видов животных?
Он должен быть у всех видов животных - согласно концепции «Анерем», если она окажется верной. Действительно, концепция исходит из всеобщности не только старения, но и мейоза. Я тщательно исследовал этот вопрос по литературным данным. Конечно, у достаточно развитых животных - у рыб и «выше» - имеется только половой способ размножения, который подразумевает и наличие мейоза. Кроме того, существуют огромные секторы и флоры и фауны, в которых распространены смешанные типы размножения. Это означает, что у них чередуются более или менее продолжительные акты бесполого размножения (например, митотические деления, спорообразование, почкование, фрагментация и т.д.) и единичные акты полового или квазиполового размножения. Существенная черта квазиполового процесса (так называемого эндомиксиса) состоит в том, что здесь тоже происходит соединение структурно одинаковых хромосом из отцовского и материнского набора (конъюгация гомологичных хромосом), хотя оно и не завершается их расхождением по разным клеткам.
Таким образом, при смешанном размножении несколько поколений организмов живут, как бы постепенно старея (подобно тому, как у более сложных животных стареют митотически делящиеся клетки), а затем половой процесс возвращает отдельные организмы к «нулевому» возрасту и обес-
печивает безбедную жизнь ещё нескольким поколениям. И наконец, считается, что ряд простых животных размножается только бесполым путём. Но в их отношении у меня остаётся некоторое сомнение: не просмотрели ли у этих организмов в длинной череде бесполых размножений чего-нибудь похожего на мейоз или эндомиксис (самооплодотворение)?
Получается, что развиваемая вами концепция ставит крест на всех мечтах продлить человеческую жизнь. Ведь обычные (неполовые) клетки обречены стареть и стареть?
Нет, крест я не ставлю. Во-первых, потому, что для нас гораздо важней не сам факт старения, а скорость этого процесса. А воздействовать на скорость старения можно многими средствами. Какие-то из них известны, какие-то (как ионы Скулачёва) - в стадии исследования, какие-то будут открыты позже.
Во-вторых, не исключено, что со временем удастся инициировать некоторые процессы мейоза и в соматических клетках - например, в стволовых и неделящихся. Я имею в виду те процессы, которые восстанавливают состояние генома: это, видимо, конъюгация гомологичных хромосом, кроссинговер или что-то более тонкое и пока неизвестное. Не вижу причин, по которым это было бы в принципе невозможно. В линиях половых клеток в мейоз вступают, в общем, такие же по структуре клетки, как и многие прочие. Причём и после конъюгации хромосом в последних сохраняется активность соответствующих генов. Однако для реализации этого проекта надо предварительно полностью определить гены, ответственные за разные аспекты мейоза, и установить способы целенаправленного воздействия на них. Это, конечно, весьма фантастический проект. Однако разве не казалось фантастикой вчера многое из того, что мы имеем сегодня?!
Мейоз (от греч.мейозис – уменьшение) – это особый тип деления эукариотических клеток, при котором после однократного удвоения ДНК клеткаделится дважды , и из одной диплоидной клетки образуются 4 гаплоидные. Состоит из 2-х последовательных делений (обозначаютсяIиII); каждое из них, подобно митозу, включает 4 фазы (профазу, метафазу, анафазу, телофазу) и цитокинез.
Фазы мейоза:
Профаза I , она сложная, делится на 5 стадий:
1. Лептонема (от греч.leptos – тонкий,nema – нить) – хромосомы спирализуются и становятся видны как тонкие нити. Каждая гомологичная хромосома уже реплицирована на 99,9% и состоит из двух сестринских хроматид, связанных между собой в районе центромеры. Содержание генетического материала –2 n 2 xp 4 c . Хромосомы с помощью белковых скоплений (прикрепительных дисков ) закреплены обоими концами на внутренней мембране ядерной оболочки. Ядерная оболочка сохраняется, ядрышко видно.
2. Зигонема (от греч.zygon – парный) – гомологичные диплоидные хромосомы устремляются друг к другу и соединяются сначала в области центромеры, а затем – по всей длине (конъюгация ). Образуютсябиваленты (от лат.bi – двойной,valens – сильный), илитетрады хроматид. Число бивалентов соответствует гаплоидному набору хромосом, содержание генетического материала можно записать как1 n 4 xp 8 c . Каждая хромосома в одном биваленте происходит либо от отца, либо от матери.Половые хромосомы располагаются около внутренней ядерной мембраны. Эта область называетсяполовым пузырьком.
Между гомологичными хромосомами в каждом биваленте образуются специализированные синаптонемальные комплексы (от греч.synapsis – связь, соединение), которые представляют собой белковые структуры. При большом увеличении в комплексе видны две параллельные белковые нити толщиной 10 нм каждая, соединенные тонкими поперечными полосами размерами около 7 нм, по обе стороны от них лежат хромосомы в виде множества петель.
В центре комплекса проходит осевой элемент толщиной 20 – 40 нм. Синаптонемальный комплекс сравнивают сверевочной лестницей , стороны которой образованы гомологичными хромосомами. Более точное сравнение –застежка типа «молния» .
К концу зигонемы каждая пара гомологичных хромосом связана между собой с помощью синаптонемальных комплексов. Лишь половые хромосомы XиYконъюгируют не полностью, т. к. они неполностью гомологичны.
3. В пахинеме (от греч.pahys – толстый) биваленты укорачиваются и утолщаются. Между хроматидами материнского и отцовского происхождения в нескольких местах возникают соединения –хиазмы (от греч.chiazma – перекрест). В области каждой хиазмы формируется комплекс белков, участвующих врекомбинации (d~ 90 нм), и происходит обмен соответствующих участков гомологичных хромосом – от отцовской к материнской и наоборот. Этот процесс называюткросссинговером (от англ.с rossing - over – перекресток). В каждом биваленте человека, например, кроссинговер происходит в двух – трех участках.
4. В диплонеме (от греч.diploos – двойной) синаптонемальные комплексы распадаются, и гомологичные хромосомы каждого бивалентаотодвигаются друг от друга , но связь между ними сохраняется в зонах хиазм.
5. Диакинез (от греч.diakinein – проходить через). В диакинезе завершается конденсация хромосом, они отделяются от ядерной оболочки, но гомологичные хромосомы продолжают еще оставаться связанными между собой концевыми участками, а сестринские хроматиды каждой хромосомы – центромерами. Биваленты приобретают причудливую формуколец, крестов, восьмерок и т. д. В это время разрушаются ядерная оболочка и ядрышки. Реплицированные центриоли направляются к полюсам, к центромерам хромосом прикрепляются нити веретена деления.
В целом профаза мейоза очень длительна. При развитии спермиев она может длиться несколько суток, а при развитии яйцеклеток – в течение многих лет.
Метафаза I напоминает аналогичную стадию митоза. Хромосомы устанавливаются в экваториальной плоскости, образуя метафазную пластинку. В отличие от митоза, микротрубочки веретена прикрепляются к центромере каждой хромосомы лишь с одной стороны (со стороны полюса), а центромеры гомологичных хромосом расположены по обеим сторонам экватора. Связь между хромосомами с помощью хиазм продолжает сохраняться.
В анафазе I хиазмы распадаются, гомологичные хромосомы отделяются друг от друга и расходятся к полюсам.Центромеры этих хромосом, однако, в отличие от анафазы митоза,не реплицируются , а значит, сестринские хроматиды не расходятся. Расхождение хромосом носитслучайный характер . Содержание генетической информации становится1 n 2 xp 4 c у каждого полюса клетки, а в целом в клетке –2(1 n 2 xp 4 c ) .
В телофазе I , как и при митозе, формируются ядерные оболочки и ядрышки, образуется и углубляетсяборозда деления. Затем происходитцитокинез . В отличие от митоза, деспирализации хромосом не происходит.
В результате мейоза Iобразуются 2 дочерние клетки, содержащие гаплоидный набор хромосом; при этом каждая хромосома имеет 2 генетически отличные (рекомбинантные) хроматиды:1 n 2 xp 4 c . Следовательно, в результате мейозаIпроисходитредукция (уменьшение вдвое) числа хромосом, откуда и название первого деления –редукционное .
После окончания мейоза Iнаступает короткий промежуток -интеркинез , в течение которого не происходит репликации ДНК и удвоения хроматид.
Профаза II недлительна, и конъюгации хромосом при этом не наступает.
В метафазе II хромосомы выстраиваются в плоскости экватора.
В анафазе II ДНК в области центромеры реплицируется, как это происходит и в анафазе митоза, хроматиды расходятся к полюсам.
Послетелофазы II ицитокинеза II образуются дочерние клетки с содержанием генетического материала в каждой –1 n 1 xp 2 c . В целом, второе деление называетсяэквационным (уравнительным).
Итак, в результате двух последовательных делений мейоза образуются 4 клетки, каждая из которых несет гаплоидный набор хромосом.
Отдельные фазы мейоза у животных описал В. Флемминг (1882), а у растений - Э.Страсбургер (1888), а затем российский ученый В.И. Беляев. В это же время (1887) А. Вайсман теоретически обосновал необходимость мейоза как механизма поддержания постоянного числа хромосом. Первое подробное описание мейоза в ооцитах кролика дал Уиниуортер (1900). Изучение мейоза продолжается до сих пор.
Биологическое значение мейоза
Биологическое значение мейоза заключается в поддержании постоянства числа хромосом при наличии полового процесса. Кроме того, вследствие кроссинговера происходит рекомбинация - появление новых сочетаний наследственных задатков в хромосомах. Мейоз обеспечивает также комбинативную изменчивость - появление новых сочетаний наследственных задатков при дальнейшем оплодотворении.
Ход мейоза находится под контролем генотипа организма, под контролем половых гормонов (у животных), фитогормонов (у растений) и множества иных факторов (например, температуры).
Мейоз (у высших растений), имеет место накануне цветения и приводит к образованию гаплоидного гаметофита, в котором позднее образуются гаметы.
О живых организмах известно, что они дышат, питаются, размножаются и погибают, в этом состоит их биологическая функция. Но за счет чего это все происходит? За счет кирпичиков - клеток, которые тоже дышат, питаются, погибают и размножаются. Но как это происходит?
О строении клеток
Дом состоит из кирпичей, блоков или бревен. Так и организм можно разделить на элементарные единицы - клетки. Все разнообразие живых существ состоит именно из них, отличие лежит лишь в их количестве и видах. Из них состоят мышцы, костная ткань, кожа, все внутренние органы - настолько сильно они различаются в своем назначении. Но вне зависимости от того, какие функции выполняет та или иная клетка, все они устроены примерно одинаково. Прежде всего, у любого "кирпичика" есть оболочка и цитоплазма с расположенными в ней органоидами. Некоторые клетки не имеют ядра, их называют прокариотическими, однако все более или менее развитые организмы состоят из эукариотических, имеющих ядро, в котором хранится генетическая информация.
Органоиды, расположенные в цитоплазме, разнообразны и интересны, они выполняют важные функции. В клетках животного происхождения выделяют эндоплазматическую сеть, рибосомы, митохондрии, комплекс Гольджи, центриоли, лизосомы и двигательные элементы. С помощью них и происходят все процессы, которые обеспечивают функционирование организма.
Жизнедеятельность клеток
Как уже было сказано, все живое питается, дышит, размножается и умирает. Это утверждение справедливо как для цельных организмов, то есть людей, животных, растений и т. д., так и для клеток. Это удивительно, но каждый "кирпичик" обладает своей собственной жизнью. За счет своих органоидов он получает и перерабатывает питательные вещества, кислород, выводит все лишнее наружу. Сама цитоплазма и эндоплазматическая сеть выполняют транспортную функцию, митохондрии отвечают в том числе за дыхание, а также обеспечение энергией. Комплекс Гольджи занимается накоплением и выводом продуктов жизнедеятельности клетки. Остальные органоиды также участвуют в сложных процессах. И на определенном этапе своего начинает делиться, то есть происходит процесс размножения. Его стоит рассмотреть более подробно.
Процесс деления клеток
Размножение - одна из стадий развития живого организма. То же относится и к клеткам. На определенном этапе жизненного цикла они входят в состояние, когда становятся готовы к размножению. просто делятся надвое, удлиняясь, а потом образовывая перегородку. Этот процесс прост и практически полностью изучен на примере палочковидных бактерий.
С все обстоит несколько сложнее. Они размножаются тремя разными способами, которые называются амитоз, митоз и мейоз. Каждый из этих путей имеет свои особенности, он присущ определенному виду клеток. Амитоз

считается самым простым, его также называют прямым бинарным делением. При нем происходит удвоение молекулы ДНК. Однако веретено деления не образуется, так что этот способ является наиболее энергетически экономичным. Амитоз наблюдается у одноклеточных организмов, в то время как ткани многоклеточных размножаются с помощью других механизмов. Однако он иногда наблюдается и там, где снижена митотическая активность, например, в зрелых тканях.
Иногда прямое деление выделяют как разновидность митоза, однако некоторые ученые считают это отдельным механизмом. Протекание этого процесса даже в старых клетках происходит довольно редко. Далее будут рассмотрены мейоз и его фазы, процесс митоза, а также сходства и различия этих способов. По сравнению с простым делением они более сложны и совершенны. Особенно это касается редукционного деления, так что характеристика фаз мейоза будет наиболее подробной.
Важную роль в делении клетки имеют центриоли - специальные органоиды, как правило, располагающиеся рядом с комплексом Гольджи. Каждая такая структура состоит из 27 микротрубочек, сгруппированных по три. Вся конструкция имеет цилиндрическую форму. Центриоли непосредственно участвуют в формировании веретена деления клетки в процессе непрямого деления, о котором речь пойдет дальше.
Митоз
Продолжительность существования клеток различается. Некоторые живут пару дней, а какие-то можно отнести к долгожителям, поскольку их полная смена происходит очень редко. И практически все эти клетки размножаются с помощью митоза. У большинства из них между периодами деления проходит в среднем 10-24 часа. Сам митоз занимает небольшой период времени - у животных примерно 0,5-1

час, а у растений около 2-3. Этот механизм обеспечивает рост клеточной популяции и воспроизводство идентичных по своему генетическому наполнению единиц. Так соблюдается преемственность поколений на элементарном уровне. При этом число хромосом остается неизменным. Именно этот механизм является наиболее распространенным вариантом репродукции эукариотических клеток.
Значение этого вида деления велико - этот процесс помогает расти и регенерировать тканям, за счет чего происходит развитие всего организма. Кроме того, именно митоз лежит в основе бесполого размножения. И еще одна функция - перемещение клеток и замена уже отживших. Поэтому считать, что из-за того, что стадии мейоза сложнее, то и его роль гораздо выше, неправильно. Оба эти процесса выполняют разные функции и по-своему важны и незаменимы.
Митоз состоит из нескольких фаз, различающихся по своим морфологическим особенностям. Состояние, в котором клетка находится, будучи готовой к непрямому делению, называют интерфазой, а непосредственно процесс разделяется еще на 5 стадий, которые необходимо рассмотреть подробнее.

Фазы митоза
Находясь в интерфазе, клетка готовится к делению: происходит синтез ДНК и белков. Эта стадия подразделяется на еще несколько, в ходе которых происходит рост всей структуры и удвоение хромосом. В этом состоянии клетка пребывает до 90% всего жизненного цикла.
Остальные 10% занимает непосредственно деление, разделяющееся на 5 стадий. При митозе клеток растений также выделяется препрофаза, которая отсутствует во всех других случаях. Происходит образование новых структур, ядро перемещается к центру. Формируется препрофазная лента, размечающая предполагаемое место будущего деления.
Во все же остальных клетках процесс митоза проходит следующим образом:
Таблица 1
| Наименование стадии | Характеристика |
| Профаза | Ядро увеличивается в размерах, хромосомы в нем спирализуются, становятся видимыми в микроскоп. В цитоплазме образуется веретено деления. Зачастую происходит распад ядрышка, однако это происходит не всегда. Содержание генетического материала в клетке остается неизменным. |
| Прометафаза | Происходит распад ядерной мембраны. Хромосомы начинают активное, но беспорядочное движение. В конечном счете, все они приходят в плоскость метафазной пластинки. Этот этап длится до 20 минут. |
| Метафаза | Хромосомы выстраиваются вдоль экваториальной плоскости веретена деления примерно на равном расстоянии от обоих полюсов. Численность микротрубочек, удерживающих всю конструкцию в стабильном состоянии, достигает максимума. Сестринские хроматиды отталкиваются друг от друга, сохраняя соединение лишь в центромере. |
| Анафаза | Наиболее короткая стадия. Хроматиды разделяются и отталкиваются друг от друга в направлении ближайших полюсов. Этот процесс иногда выделяют отдельно и называют анафазой А. В дальнейшем происходит расхождение самих полюсов деления. В клетках некоторых простейших веретено деления при этом увеличивается в длину до 15 раз. И этот подэтап носит название анафаза В. Длительность и последовательность процессов на данной стадии вариабельна. |
| Телофаза | После окончания расхождения к противоположным полюсам хроматиды останавливаются. Происходит деконденсация хромосом, то есть их увеличение в размерах. Начинается реконструкция ядерных оболочек будущих дочерних клеток. Микротрубочки веретена деления исчезают. Формируются ядра, возобновляется синтез РНК. |
После завершения деления генетической информации происходит цитокинез или цитотомия. Под этим термином подразумевается образование тел дочерних клеток из тела материнской. При этом органоиды, как правило, делятся пополам, хотя возможны исключения, образуется перегородка. Цитокинез не выделяют в отдельную фазу, как правило, рассматривая его в рамках телофазы.
Итак, в самых интересных процессах задействованы хромосомы, которые несут генетическую информацию. Что же это такое и почему они так важны?
О хромосомах
Еще не имея ни малейшего понятия о генетике, люди знали, что многие качества потомства зависят от родителей. С развитием биологии стало очевидно, что информация о том или ином организме хранится в каждой клетке, и часть ее передается будущим поколениям.
В конце 19 века были открыты хромосомы - структуры, состоящие из длинной

молекулы ДНК. Это стало возможно с совершенствованием микроскопов, и даже сейчас рассмотреть их можно лишь в период деления. Чаще всего открытие приписывают немецкому ученому В. Флемингу, который не только упорядочил все то, что было изучено до него, но и внес свой вклад: он одним из первых исследовал клеточную структуру, мейоз и его фазы, а также ввел термин "митоз". Само понятие "хромосома" было предложено чуть позже другим ученым - немецким гистологом Г. Вальдейером.
Структура хромосом в момент, когда они четко видны, довольно проста - они представляют собой две хроматиды, соединенные посередине центромерой. Она является специфической последовательностью нуклеотидов и играет важную роль в процессе размножения клеток. В конечном итоге хромосома внешне в профазе и метафазе, когда ее можно лучше всего разглядеть, напоминается букву Х.
В 1900 году были открыты описывающие принципы передачи наследственных признаков. Тогда стало окончательно ясно, что хромосомы - это именно то, с помощью чего передается генетическая информация. В дальнейшем учеными был проведен ряд экспериментов, доказывающих это. И тогда предметом изучения стало и то влияние, котрое на них оказывает деление клетки.
Мейоз
В отличие от митоза этот механизм в итоге приводит к образованию двух клеток с набором хромосом в 2 раза меньше исходного. Таким образом процесс мейоза служит переходом от диплоидной фазы к гаплоидной, причем в первую очередь

речь идет о делении ядра, а уже во вторую - всей клетки. Восстановление же полного набора хромосом происходит в результате дальнейшего слияния гамет. В связи с уменьшением количества хромосом этот метод еще определяют как редукционное деление клетки.
Мейоз и его фазы изучали такие известные ученые, как В. Флеминг, Э. Страсбургрер, В. И. Беляев и другие. Исследование этого процесса в клетках как растений, так и животных, продолжается до сих пор - настолько он сложен. Изначально этот процесс считался вариантом митоза, однако практически сразу после открытия он все-таки был выделен как отдельный механизм. Характеристика мейоза и его теоретическое значение были впервые в достаточной степени описаны Августом Вайсманом еще в 1887 году. С тех пор изучение процесса редукционного деления сильно продвинулось, но сделанные выводы пока не были опровергнуты.
Мейоз не следует путать с гаметогенезом, хотя оба эти процесса тесно связаны. В образовании половых клеток участвуют оба механизма, однако между ними есть ряд серьезных отличий. Мейоз происходит в две стадии деления, каждая из которых состоит из 4 основных фаз, между ними есть короткий перерыв. Длительность всего процесса зависит от количества ДНК в ядре и структуры хромосомной организации. В целом он гораздо более продолжителен в сравнении с митозом.
Кстати, одна из основных причин значительного видового разнообразия - именно мейоз. Набор хромосом в результате редукционного деления разбивается надвое, так что появляются новые комбинации генов, в первую очередь потенциально повышающие приспособляемость и адаптивность организмов, в итоге получающих те или иные наборы признаков и качеств.
Фазы мейоза
Как уже было упомянуто, редукционное клеточное деление условно делят на две стадии. Каждая из этих стадий разделена еще на 4. И первая фаза мейоза - профаза I в свою очередь подразделяется еще на 5 отдельных этапов. Поскольку изучение этого процесса продолжается, в дальнейшем могут быть выделены и другие. Сейчас же различают следующие фазы мейоза:
Таблица 2
| Наименование стадии | Характеристика |
| Первое деление (редукционное) | |
Профаза I | |
| лептотена | По-другому этот этап называют стадией тонких нитей. Хромосомы выглядят в микроскопе как спутанный клубок. Иногда выделяют пролептотену, когда отдельные ниточки еще сложно разглядеть. |
| зиготена | Стадия сливающихся нитей. Гомологичные, то есть сходные между собой по морфологии и в генетическом отношении, пары хромосом сливаются. В процессе слияния, то есть конъюгации, образуются биваленты, или тетрады. Так называют довольно устойчивые комплексы из пар хромосом. |
| пахитена | Стадия толстых нитей. На этом этапе хромосомы спирализуются и завершается репликация ДНК, образуются хиазмы - точки контакта отдельных частей хромосом - хроматид. Происходит процесс кроссинговера. Хромосомы перекрещиваются и обмениваются некоторыми участками генетической информации. |
| диплотена | Также называется стадией двойных нитей. Гомологичные хромосомы в бивалентах отталкиваются друг от друга и остаются связанными только в хиазмах. |
| диакинез | На этой стадии биваленты расходятся на периферии ядра. |
| Метафаза I | Оболочка ядра разрушается, формируется веретено деления. Биваленты перемещаются к центру клетки и выстраиваются вдоль экваториальной плоскости. |
| Анафаза I | Биваленты распадаются, после чего каждая хромосома из пары перемещается к ближайшему полюсу клетки. Разделения на хроматиды не происходит. |
| Телофаза I | Завершается процесс расхождения хромосом. Происходит формирование отдельных ядер дочерних клеток, каждое - с гаплоидным набором. Хромосомы деспирализуются, образуется ядерная оболочка. Иногда наблюдается цитокинез, то есть деление самого тела клетки. |
| Второе деление (эквационное) | |
| Профаза II | Происходит конденсация хромосом, клеточный центр делится. Разрушается ядерная оболочка. Образуется веретено деления, перпендикулярное первому. |
| Метафаза II | В каждой из дочерних клеток хромосомы выстраиваются вдоль экватора. Каждая из них состоит из двух хроматид. |
| Анафаза II | Каждая хромосома делится на хроматиды. Эти части расходятся к противоположным полюсам. |
| Телофаза II | Полученные однохроматидные хромосомы деспирализуются. Образуется ядерная оболочка. |
Итак, очевидно, что фазы деления мейоза гораздо сложнее, чем процесс митоза. Но, как уже было упомянуто, это не умаляет биологической роли непрямого деления, поскольку они выполняют разные функции.
Кстати, мейоз и его фазы наблюдаются и у некоторых простейших. Однако, как правило, он включает в себя лишь одно деление. Предполагается, что такая одноступенчатая форма позднее развилась в современную, двухступенчатую.
Отличия и сходства митоза и мейоза
На первый взгляд кажется, что различия двух этих процессов очевидны, ведь это совершенно разные механизмы. Однако при более глубоком анализе оказывается, что различия митоза и мейоза не так уж глобальны, в конце концов они приводят к образованию новых клеток.
Прежде всего стоит поговорить о том, что есть общего у этих механизмов. По сути совпадения всего два: в одинаковой последовательности фаз, а также в том, что

перед обоими видами деления происходит репликация ДНК. Хотя, что касается мейоза, до начала профазы I этот процесс не завершается полностью, заканчиваясь на одной из первых подстадий. А последовательность фаз хоть и аналогична, но, по сути, происходящие в них события совпадают не полностью. Так что сходства митоза и мейоза не так уж и многочисленны.
Различий же гораздо больше. Прежде всего, митоз происходит в в то время как мейоз тесно связан с образованием половых клеток и спорогенезом. В самих фазах процессы не полностью совпадают. Например, кроссинговер в митозе происходит во время интерфазы, и то не всегда. Во втором же случае на этот процесс приходится анафаза мейоза. Рекомбинация генов в непрямом делении обычно не осуществляется, а значит, он не играет никакой роли в эволюционном развитии организма и поддержании внутривидового разнообразия. Количество получившихся в результате митоза клеток - две, и они в генетическом смысле идентичны материнской и обладают диплоидным набором хромосом. Во время редукционного деления все иначе. Результат мейоза - 4 отличающихся от материнской. Кроме того, оба механизма значительно различаются по длительности, и это связано не только с различием в количестве ступеней деления, но и длительностью каждого из этапов. Например, первая профаза мейоза длится намного дольше, ведь в это время происходит конъюгация хромосом и кроссинговер. Именно поэтому ее дополнительно делят на несколько стадий.
В общем и целом сходства митоза и мейоза достаточно незначительны по сравнению с их отличиями друг от друга. Перепутать эти процессы практически невозможно. Поэтому сейчас даже несколько удивляет то, что редукционное деление раньше считалось разновидностью митоза.
Последствия мейоза
Как уже было упомянуто, после окончания процесса редукционного деления, вместо материнской клетки с диплоидным набором хромосом образуются четыре гаплоидных. И если говорить про различия митоза и мейоза - это самое значительное. Восстановление необходимого количества, если речь идет о половых клетках, происходит после оплодотворения. Таким образом, с каждым новым поколением не происходит удвоения количества хромосом.
Кроме того, во время мейоза происходит В процессе размножения это приводит к поддержанию внутривидового разнообразия. Так что тот факт, что даже родные братья и сестры порой сильно отличаются друг от друга - именно результат мейоза.
Кстати, стерильность некоторых гибридов в животном мире - тоже проблема редукционного деления. Дело в том, что хромосомы родителей, принадлежащих к разным видам, не могут вступить в конъюгацию, а значит, процесс образования полноценных жизнеспособных половых клеток невозможен. Таким образом, именно мейоз лежит в основе эволюционного развития животных, растений и других организмов.
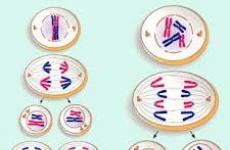
Деление клеток посредством мейоза проходит в два основных этапа: мейоз I и мейоз II. В конце мейотического процесса образуются четыре . Прежде чем делящаяся клетка попадет в мейоз, она проходит через период , называемый интерфазой.
Интерфаза
- Фаза G1: этап развития клетки перед синтезом ДНК. На этой стадии клетка подготавливаясь к делению увеличивается в массе.
- S-фаза: период, в течение которого синтезируется ДНК. Для большинства клеток эта фаза занимает короткий промежуток времени.
- Фаза G2: период после синтеза ДНК, но до начала профазы. Клетка продолжает синтезировать дополнительные белки и увеличиваться в размерах.
В последней фазе интерфазы клетка все еще имеет нуклеолы. окружено ядерной мембраной, а клеточное хромосомы дублируются, но находятся в форме . В две пары , образованных из репликации одной пары, расположены за пределами ядра. В конце интерфазы клетка переходит в первый этап мейоза.
Мейоз I:
Профаза I
В профазе I мейоза происходят следующие изменения:
- Хромосомы конденсируются и присоединяются к ядерной оболочке.
- Возникает синапсис (попарное сближение гомологичных хромосом) и образуется тетрада. Каждая тетрада состоит из четырех хроматид.
- Может произойти генетическая рекомбинация.
- Хромосомы сгущаются и отсоединяются от ядерной оболочки.
- Подобно , центриоли мигрируют друг от друга, а ядерная оболочка и ядрышки разрушаются.
- Хромосомы начинают миграцию к метафазной (экваториальной) пластине.
В конце профазы I клетка входит в метафазу I.
Метафаза I
 В метафазе I мейоза происходят следующие изменения:
В метафазе I мейоза происходят следующие изменения:
- Тетрады выравниваются на метафазной пластине.
- гомологичных хромосом ориентированы на противоположные полюса клетки.
В конце метафазы I клетка входит в анафазу I.
Анафаза I
 В анафазе I мейоза происходят происходят следующие изменения:
В анафазе I мейоза происходят происходят следующие изменения:
- Хромосомы перемещаются в противоположные концы клетки. Подобно митозу, кинетохоры взаимодействуют с микротрубочками, чтобы переместить хромосомы к полюсам клетки.
- В отличие от митоза, остаются вместе после того, как перемещаются в противоположные полюса.
В конце анафазы I клетка входит в телофазу I.
Телофаза I
 В телофазе I мейоза происходят следующие изменения:
В телофазе I мейоза происходят следующие изменения:
- Волокна веретена продолжают перемещать гомологичные хромосомы на полюса.
- Как только движение завершено, каждый полюс клетки имеет гаплоидное количество хромосом.
- В большинстве случаев цитокинез (деление ) происходит одновременно с телофазой I.
- В конце телофазы I и цитокинеза образуются две дочерние клетки, каждая из которых имеет половину числа хромосом исходной родительской клетки.
- В зависимости от типа клетки могут возникать различные процессы при подготовке к мейозу II. Однако генетический материал не реплицируется снова.
В конце телофазы I клетка входит в профазу II.
Мейоз II:
Профаза II
 В профазе II мейоза происходят следующие изменения:
В профазе II мейоза происходят следующие изменения:
- Ядерная и ядра разрушаются, пока появляется веретено деления.
- Хромосомы больше не реплицируются в этой фазе.
- Хромосомы начинают мигрировать к метафазной пластинке II (на экваторе клеток).
В конце профазы II клетки входят в метафазу II.
Метафаза II
 В метафазе II мейоза происходят следующие изменения:
В метафазе II мейоза происходят следующие изменения:
- Хромосомы выстраиваются на метафазной пластинке II в центре клеток.
- Кинетохорные нити сестринских хроматид расходятся к противоположным полюсам.
В конце метафазы II клетки входят в анафазу II.
Анафаза II
 В анафазе II мейоза происходят следующие изменения:
В анафазе II мейоза происходят следующие изменения:
- Сестринские хроматиды разделяются и начинают перемещаться к противоположным концам (полюсам) клетки. Волокна веретена деления, не связанные с хроматидами, вытягиваются и удлиняют клетки.
- Как только парные сестринские хроматиды отделены друг от друга, каждая из них считается полной хромосомой, называемые .
- При подготовке к следующему этапу мейоза два полюса клеток также отдаляются друг от друга во время анафазы II. В конце анафазы II каждый полюс содержит полную компиляцию хромосом.
После анафазы II клетки входят в телофазу II.
Телофаза II
 В телофазе II мейоза происходят следующие изменения:
В телофазе II мейоза происходят следующие изменения:
- Образуются отдельные ядра на противоположных полюсах.
- Происходит цитокинез (деление цитоплазмы и образование новых клеток).
- В конце мейоза II производятся четыре дочерние клетки. Каждая клетка имеет половину числа хромосом от исходной родительской клетки.
Результат мейоза
Конечным результатом мейоза является производство четырех дочерних клеток. Эти клетки имеют в двое меньше хромосом относительно родительской. При мейозе продуцируются только половые . Другие делятся посредством митоза. Когда половые объединяются во время оплодотворения, они становятся . Диплоидные клетки имеют полный набор гомологичных хромосом.