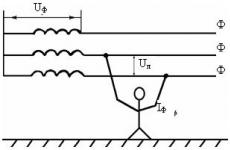
Transporte de gases no corpo. Características do sistema respiratório humano
A troca gasosa de 02 e CO2 através da membrana alvéolo-capilar ocorre com a ajuda de difusão, que é realizado em duas etapas. No primeiro estágio, a transferência de gases por difusão ocorre através da barreira ar-sangue; no segundo estágio, os gases se ligam ao sangue dos capilares pulmonares, cujo volume é de 80-150 ml com a espessura da camada sanguínea nos capilares apenas 5-8 mícrons. O plasma sanguíneo praticamente não impede a difusão dos gases, ao contrário da membrana eritrocitária.
A estrutura dos pulmões cria condições fávoraveis para troca gasosa: a zona respiratória de cada pulmão contém cerca de 300 milhões de alvéolos e aproximadamente o mesmo número de capilares, tem uma área de 40-140 m 2 com uma espessura da barreira ar-sangue de apenas 0,3-1,2 mícrons.
As características da difusão gasosa são caracterizadas quantitativamente através capacidade de difusão dos pulmões. Para 02, a capacidade de difusão dos pulmões é o volume de gás transferido dos alvéolos para o sangue em 1 minuto com um gradiente de pressão de gás alvéolo-capilar de 1 mm Hg.
O movimento dos gases ocorre como resultado da diferença nas pressões parciais. Pressão parcial é a parte da pressão que um determinado gás constitui a partir da mistura total de gases. A pressão reduzida 0n no tecido promove o movimento de oxigênio para ele. Para CO2, o gradiente de pressão é direcionado para lado reverso, e o CO com o ar exalado vai para o meio ambiente. O estudo da fisiologia da respiração resume-se realmente ao estudo desses gradientes e como eles são mantidos.
O gradiente de pressão parcial de oxigênio e dióxido de carbono é a força com a qual as moléculas desses gases tendem a penetrar através da membrana alveolar no sangue. A tensão parcial de um gás no sangue ou nos tecidos é a força com que as moléculas de um gás solúvel tendem a escapar para um meio gasoso.
Ao nível do mar, a pressão atmosférica é em média 760 mm Hg e a percentagem de oxigénio é de cerca de 21%. Neste caso, p02 na atmosfera é: 760 x 21/100 = 159 mm Hg. Ao calcular a pressão parcial dos gases no ar alveolar, deve-se levar em consideração que este ar contém vapor d’água (47 mm Hg). Portanto, este número é subtraído do valor
pressão atmosférica, e a fração da pressão parcial dos gases é responsável por (760 - 47) == 713 mm Hg. Com um teor de oxigênio no ar alveolar igual a 14%, sua pressão parcial será de 100 mm Hg. Arte. Com um teor de dióxido de carbono de 5,5%, a pressão parcial será de cerca de 40 mm Hg.
No sangue arterial, a pressão parcial de oxigênio atinge quase 100 mm Hg, no sangue venoso - cerca de 40 mm Hg, e no fluido tecidual, nas células - 10 - 15 mm Hg. A tensão do dióxido de carbono no sangue arterial é de cerca de 40 mm Hg, no sangue venoso - 46 mm Hg e nos tecidos - até 60 mm Hg.
Os gases no sangue estão em dois estados: fisicamente dissolvidos e quimicamente ligados. A dissolução ocorre de acordo com a lei de Henry, segundo a qual a quantidade de gás dissolvido em um líquido é diretamente proporcional à pressão parcial desse gás sobre o líquido. Para cada unidade de pressão parcial, 0,003 ml de 02, ou 3 ml/l de sangue, são dissolvidos em 100 ml de sangue.
Cada gás tem seu próprio coeficiente de solubilidade. À temperatura corporal, a solubilidade do CO2 é 25 vezes maior que o 02. Devido à boa solubilidade do dióxido de carbono no sangue e nos tecidos, o CO2 é transferido 20 vezes mais facilmente que o 02. A tendência de um gás passar de um líquido para um a fase gasosa é chamada de tensão do gás. Em condições normais, em 100 ml de sangue, apenas 0,3 ml de 02 e 2,6 ml de CO2 estão dissolvidos. Tais valores não conseguem atender às demandas do organismo em 02.
A troca gasosa de oxigênio entre o ar alveolar e o sangue ocorre devido à presença de um gradiente de concentração 02 entre esses meios. O transporte de oxigênio começa nos capilares dos pulmões, onde a maior parte do O2 que entra no sangue entra em uma ligação química com a hemoglobina. A hemoglobina é capaz de se ligar seletivamente ao O2 e formar oxiemoglobina (HbO2). Um grama de hemoglobina liga 1,36 - 1,34 ml SOBRE 2a 1 litro de sangue contém 140-150 g de hemoglobina. Existem 1,39 ml de oxigênio por 1 grama de hemoglobina. Conseqüentemente, em cada litro de sangue, o conteúdo máximo possível de oxigênio na forma quimicamente ligada será de 190 - 200 ml 02, ou 19 vol% - isto é capacidade de oxigênio do sangue. O sangue humano contém aproximadamente 700-800 g de hemoglobina e pode ligar 1 litro de oxigênio.
A capacidade de oxigênio do sangue é a quantidade SOBRE 2, que é ligado ao sangue até que a hemoglobina esteja completamente saturada. Uma alteração na concentração de hemoglobina no sangue, por exemplo, na anemia, envenenamento por venenos, altera sua capacidade de oxigênio. Ao nascer, a pessoa apresenta valores mais elevados de capacidade de oxigênio e concentração de hemoglobina no sangue. A saturação de oxigênio no sangue expressa a relação entre a quantidade de oxigênio ligado e a capacidade de oxigênio do sangue, ou seja, sob saturação sanguínea 0^
refere-se à porcentagem de oxiemoglobina em relação à hemoglobina presente no sangue. Em condições normais, a saturação de 0^ é 95-97%. Ao respirar com oxigênio puro, a saturação sanguínea O^ atinge 100%, e ao respirar com uma mistura gasosa com baixo teor de oxigênio, o percentual de saturação cai. Em 60-65% ocorre perda de consciência.
A dependência da ligação do oxigênio pelo sangue em sua pressão parcial pode ser representada como um gráfico, onde p02 no sangue é plotado ao longo da abcissa e a saturação de oxigênio da hemoglobina é plotada ao longo da ordenada. Este gráfico é curva de dissociação da oxihemoglobina, ou curva de saturação, mostra qual proporção de hemoglobina em um determinado sangue está associada a 02 em uma ou outra de suas pressões parciais, e o que está dissociado, ou seja, livre de oxigênio. A curva de dissociação tem formato de S. O platô da curva é característico do sangue arterial saturado 02 (saturado), e a parte descendente íngreme da curva é característica do sangue venoso ou dessaturado nos tecidos (Fig. 21).
Arroz. 21. Curvas de dissociação da oxihemoglobina no sangue total em várias alterações de pH (L) e temperatura do sangue (B)
Curvas 1-6 correspondem a 0°, 10°, 20°, 30°, 38° e 43°C
A afinidade do oxigênio pela hemoglobina e a capacidade de doar 02 aos tecidos dependem das necessidades metabólicas das células do corpo e são reguladas pelos fatores mais importantes do metabolismo tecidual, causando um deslocamento na curva de dissociação. Esses fatores incluem: a concentração de íons de hidrogênio, temperatura, tensão parcial de dióxido de carbono e o composto que se acumula nos eritrócitos é o fosfato de 2,3-difosfoglicerato (DPG).Uma diminuição no pH do sangue causa um deslocamento na curva de dissociação para a direita , e um aumento no pH do sangue causa um deslocamento da curva para a esquerda. Devido ao aumento do conteúdo de CO2 nos tecidos, o pH também é mais baixo do que no plasma sanguíneo.O valor do pH e o conteúdo de CO2 nos tecidos do corpo alteram a afinidade da hemoglobina para SOBRE 2 Sua influência na curva de dissociação da oxiemoglobina é chamada de efeito Bohr (H. Bohr, 1904). Com o aumento da concentração de íons hidrogênio e da voltagem parcial de CO2 no meio, a afinidade da hemoglobina pelo oxigênio diminui. Esse “efeito” tem um importante valor adaptativo: o CO2 dos tecidos entra nos capilares, então o sangue, com o mesmo pO2, consegue liberar mais oxigênio. O metabólito 2,3-DFG formado durante a quebra da glicose também reduz a afinidade da hemoglobina pelo oxigênio.
A temperatura também influencia a curva de dissociação da oxiemoglobina. Um aumento na temperatura aumenta significativamente a taxa de degradação da oxiemoglobina e reduz a afinidade da hemoglobina para SOBRE 2 Um aumento na temperatura nos músculos em atividade promove a liberação SOBRE 2 A ligação do 02 pela hemoglobina reduz a afinidade de seus grupos amino pelo CO2 (efeito Holden). A difusão do CO2 do sangue para os alvéolos é proporcionada pela ingestão de CO2 dissolvido no plasma sanguíneo (5-10%), dos bicarbonatos (80-90%) e, finalmente, dos compostos carbâmicos dos eritrócitos (5-15 %), que são capazes de se dissociar.
O dióxido de carbono no sangue está em três frações: fisicamente dissolvido, quimicamente ligado na forma de bicarbonatos e quimicamente ligado à hemoglobina na forma de carbohemoglobina. O sangue venoso contém apenas 580 ml de dióxido de carbono. Ao mesmo tempo, a proporção de gás fisicamente dissolvido é de 25 ml, a proporção de carbohemoglobina é de cerca de 45 ml, a proporção de bicarbonatos é de 510 ml (bicarbonatos plasmáticos - 340 ml, eritrócitos - 170 ml). O sangue arterial contém menos ácido carbônico.
O processo de ligação do CO2 pelo sangue depende da tensão parcial do dióxido de carbono fisicamente dissolvido. O dióxido de carbono entra no eritrócito, onde existe uma enzima anidrase carbônica, o que pode aumentar a taxa de formação de ácido carbônico em 10.000 vezes. Depois de passar pelo eritrócito, o ácido carbônico é convertido em bicarbonato e transportado para os pulmões.
Os glóbulos vermelhos transportam 3 vezes mais CO2 que o plasma. As proteínas plasmáticas são 8 g por 100 cm 3 de sangue, enquanto a hemoglobina está contida no sangue 15 g por 100 cm 3. A maior parte do CO2 é transportada no corpo em estado ligado na forma de bicarbonatos e compostos carbâmicos, o que aumenta o tempo de troca de CO2.
Além do CO2 molecular fisicamente dissolvido no plasma sanguíneo, ele se difunde do sangue para os alvéolos dos pulmões. ENTÃO 2, que é liberado dos compostos carbâmicos dos eritrócitos devido à reação de oxidação da hemoglobina nos capilares do pulmão, bem como dos bicarbonatos plasmáticos como resultado de sua rápida
um enxame de dissociação com a ajuda da enzima anidrase carbônica contida nos eritrócitos. Esta enzima está ausente no plasma. Os bicarbonatos plasmáticos para liberar CO2 devem primeiro entrar nos eritrócitos para serem expostos à anidrase carbônica. O plasma contém bicarbonato de sódio e os eritrócitos contêm bicarbonato de potássio. A membrana dos eritrócitos é bem permeável ao CO2, portanto, parte do CO2 se difunde rapidamente do plasma para os eritrócitos. A maior quantidade de bicarbonatos plasmáticos é formada com a participação da anidrase carbônica eritrocitária.
Deve-se notar que o processo de remoção do CO2 do sangue para os alvéolos do pulmão é menos limitado do que a oxigenação do sangue, uma vez que o CO2 molecular penetra através das membranas biológicas mais facilmente do que o Ox.
Vários venenos que limitam o transporte de 0^, como CO, nitritos, ferrocianetos e muitos outros, praticamente não têm efeito no transporte de CO2. Os bloqueadores da anidrase carbônica também nunca interrompem completamente a formação de CO2 molecular. E, finalmente, os tecidos têm uma grande capacidade tampão, mas não estão protegidos contra deficiências. SOBRE 2 A excreção de CO2 pelos pulmões pode ser prejudicada com uma diminuição significativa da ventilação pulmonar (hipoventilação) como resultado de doença pulmonar, doença do trato respiratório, intoxicação ou desregulação da respiração. A retenção de CO2 leva à acidose respiratória - uma diminuição na concentração de bicarbonatos, uma mudança no pH do sangue para o lado ácido. A remoção excessiva de CO2 durante a hiperventilação durante o trabalho muscular intenso, ao subir a grandes altitudes, pode causar alcalose respiratória, uma mudança no pH do sangue para o lado alcalino.
Ao estudar a respiração externa, são utilizados os seguintes conceitos:
Ar alveolar- contido nos alvéolos após uma expiração normal;
Ar exalado- as primeiras porções de ar exalado são uma mistura de ar alveolar e ar do espaço morto.
Composição do ar em %
Como resultado das trocas gasosas entre o sangue e o ar alveolar, o sangue venoso é convertido em sangue arterial.
Fatores que determinam a difusão de gases nos pulmões.
I Gradiente alvéolo-capilar.
II Relação entre ventilação e perfusão.
III Comprimento do trajeto de perfusão.
IV Capacidade de difusão dos gases.
V Área de difusão.
1) A diferença entre pressão parcial e tensão.
A pressão parcial é a parte da pressão de uma mistura de gases por parcela de um gás.
A pressão parcial depende de:
b) do valor da pressão total: Calculada conforme a seguinte fórmula.
Por exemplo, O 2 no ar atmosférico
100% gás - 760 mmHg Arte.
x = 159 mmHg Arte. no ar atmosférico.
Ao calcular a pressão parcial do gás no ar alveolar, é necessário levar em consideração a pressão do vapor d'água ali localizado = 47 mm Hg. Arte.
Pressão parcial do gásé a força com que um gás dissolvido em um líquido tende a sair dele. Normalmente, um equilíbrio dinâmico é estabelecido entre o gás no líquido e acima do líquido.
Na circulação pulmonar, o O 2 vai para o sangue venoso vindo dos pulmões e o CO 2 do sangue vai para os pulmões.
A força motriz é o gradiente alvéolo-capilar.
Para O 2 AKG = 60 mm Hg. Art., para CO 2 - 6 mm Hg. Arte. Aqueles. as propriedades de difusão do CO2 são superiores às do O 2 .
2) A relação entre ventilação e perfusão\u003d MAV / IOC \u003d 4 - 6 / 4,5 - 5 \u003d 0,8 - 1,1 - normal.
A ventilação e a perfusão pulmonar devem corresponder. No entanto, a distribuição do fluxo sanguíneo através dos pulmões em humanos não é uniforme. Depende da posição do corpo e das mudanças sob a influência da gravidade. Na posição vertical, o valor Q do fluxo sanguíneo por unidade de volume de tecido diminui quase linearmente de baixo para cima, e a parte superior dos pulmões recebe menos sangue. Deitado, o fluxo sanguíneo aumenta no ápice, mas não muda na base. No entanto, deitado de costas, na parte posterior dos pulmões, o fluxo sanguíneo é maior do que na frente.
Durante o trabalho, o fluxo sanguíneo é aproximadamente o mesmo em todos os departamentos.
A posição vertical também afeta a ventilação. Sua intensidade aumenta de cima para baixo (assim como o fluxo sanguíneo).
No entanto, o VPO não é uniforme nos diferentes departamentos.
Os mecanismos que adaptam o fluxo sanguíneo à ventilação são reações vasomotoras e broncomotoras às mudanças na composição gasosa do ar alveolar.
Vasoconstrição com diminuição da pO 2 nos alvéolos, ou RCO 2.
broncoconstrição em ↓ РСО 2 no ar alveolar.
VPO é afetado por:
a) ventilação desigual dos pulmões nas diferentes posições do corpo no espaço;
b) a natureza do fluxo sanguíneo pulmonar dependendo da posição do corpo e da atividade do corpo;
c) velocidade do fluxo sanguíneo
3) Comprimento do caminho.
CO2; O 2 passa pelo caminho: parede alveolar + espaço intercelular + membrana basal capilar + endotélio capilar + camada plasmática + membrana eritrocitária. Um aumento na extensão do caminho - uma deterioração na oxigenação do sangue - relação inversa.
5) Área de difusão- depende da superfície dos alvéolos e capilares através dos quais ocorre a difusão (dependência direta).
A respiração é a função mais importante do corpo, pois garante a manutenção de um nível ideal de processos redox nas células, a respiração celular.
Órgãos especializados (nariz, pulmões, diafragma, coração) e células (eritrócitos, células nervosas, quimiorreceptores de vasos sanguíneos e células nervosas do cérebro que formam o centro respiratório) estão envolvidos na garantia do processo respiratório.
Convencionalmente, o processo respiratório pode ser dividido em três etapas principais: respiração externa, transporte de gases (oxigênio e dióxido de carbono) pelo sangue (entre os pulmões e as células) e respiração tecidual (oxidação de diversas substâncias nas células).
respiração externa- troca gasosa entre o corpo e o ar atmosférico circundante.
Transporte de gases pelo sangue. O principal transportador de oxigênio é a hemoglobina, uma proteína encontrada dentro dos glóbulos vermelhos. Com a ajuda da hemoglobina, até 20% do dióxido de carbono também é transportado.
Tecido ou respiração interna. Este processo pode ser condicionalmente dividido em dois: a troca de gases entre o sangue e os tecidos, o consumo de oxigênio pelas células e a liberação de dióxido de carbono (respiração intracelular, endógena).
Obviamente, o estado de saúde é determinado pelo estado da função respiratória, e pela capacidade de reserva do corpo, a reserva de saúde depende da capacidade de reserva do sistema respiratório.
Transporte de gases pelo sangue
No corpo, o oxigênio e dióxido de carbono transportado pelo sangue. O oxigênio que vem do ar alveolar para o sangue liga-se à hemoglobina dos eritrócitos, formando a chamada oxiemoglobina, e nesta forma é entregue aos tecidos. Nos capilares dos tecidos, o oxigênio é dividido e passa para os tecidos, onde é incluído nos processos oxidativos. A hemoglobina livre liga-se ao hidrogênio e se transforma na chamada hemoglobina reduzida. O dióxido de carbono formado nos tecidos passa para o sangue e entra nos glóbulos vermelhos. Então parte do dióxido de carbono combina-se com a hemoglobina reduzida, formando a chamada carbhemoglobina, e nesta forma o dióxido de carbono é entregue aos pulmões. Porém, a maior parte do dióxido de carbono dos eritrócitos, com a participação da enzima anidrase carbônica, é convertida em bicarbonatos, que passam para o plasma e são transportados para os pulmões. Nos capilares pulmonares, os bicarbonatos são decompostos com a ajuda de uma enzima especial e o dióxido de carbono é liberado. O dióxido de carbono é separado da hemoglobina. O dióxido de carbono passa para o ar alveolar e é removido com o ar exalado para o ambiente externo.
3…. Características do processo de proteção do corpo contra a influência de fatores externos e internos. Mecanismos de defesa congênitos: reflexos protetores incondicionados, funções de barreira da pele e membranas mucosas, síndrome de adaptação
A pele está intimamente ligada a todos os órgãos e sistemas do corpo. Desempenha muitas funções importantes, sendo as principais as protetoras, respiratórias, de absorção, excretoras e formadoras de pigmentos. Além disso, a pele participa de reações vasculares, termorregulação, processos metabólicos, reações neuro-reflexas do corpo.
Função protetora a pele é muito diversificada. A proteção mecânica contra estímulos externos é fornecida por um estrato córneo denso, especialmente nas palmas das mãos e plantas dos pés. Devido a essas propriedades, a pele é capaz de resistir a influências mecânicas - pressão, hematomas, rasgos, etc.
A pele protege amplamente o corpo da exposição à radiação. Os raios infravermelhos são quase completamente bloqueados pelo estrato córneo, ultravioleta - parcialmente. Penetrando profundamente na epiderme, os raios UV estimulam a produção de pigmento - melanina absorvendo os raios UV e protegendo assim as células dos efeitos nocivos do excesso de radiação e da insolação (exposição à radiação solar).
Em defesa contra irritantes químicos a queratina do estrato córneo desempenha um papel importante. A principal barreira à penetração de eletrólitos, não eletrólitos e água na pele é a camada transparente e a parte mais profunda do estrato córneo, rica em colesterol.
Proteção contra microorganismos fornecido com propriedades bactericidas da pele. O número de diferentes microrganismos na superfície da pele humana saudável varia de 115 mil a 32 milhões por 1 cm quadrado. A pele intacta é impermeável a microorganismos.
adaptativo síndrome - um conjunto de reações protetoras do corpo humano ou animal (principalmente do sistema endócrino) sob estresse. Na síndrome de adaptação, os estágios de ansiedade (mobilização de forças protetoras), resistência (adaptação a situação difícil), exaustão (com estresse severo e prolongado pode terminar em morte). Os conceitos de síndrome de adaptação e estresse foram apresentados por G. Selye.
Existem três estágios no desenvolvimento da síndrome de adaptação:
Estágio ansiedade: dura de várias horas a dois dias. Inclui duas fases - choque e contra-choque (nesta última são mobilizadas as reações de defesa do organismo).
No palco resistência a resistência do corpo a várias influências aumenta. O segundo estágio leva à estabilização ou é substituído pelo último estágio - exaustão.
Estágio exaustão: as reações de defesa estão enfraquecidas, o próprio corpo e a psique estão cansados.
A síndrome de adaptação também apresenta sinais fisiológicos: aumento do córtex adrenal, diminuição do timo, baço e gânglios linfáticos, distúrbios metabólicos com predomínio de processos de decomposição.
BILHETE 27
Ciclo de atividade cardíaca
O trabalho mecânico do coração está associado à contração do miocárdio. O trabalho do ventrículo direito é três vezes menor que o trabalho do ventrículo esquerdo.
Do ponto de vista mecânico, o coração é uma bomba de ação rítmica, facilitada pelo aparelho valvar. As contrações e relaxamentos rítmicos do coração proporcionam um fluxo contínuo de sangue. A contração do músculo cardíaco é chamada sístole, seu relaxamento - diástole. A cada sístole ventricular, o sangue é ejetado do coração para a aorta e para o tronco pulmonar.
Em condições normais, a sístole e a diástole são claramente coordenadas no tempo. O período, incluindo uma contração e subsequente relaxamento do coração, é ciclo cardíaco. Sua duração em um adulto é de 0,8 segundos com frequência de contrações de 70 a 75 vezes por minuto. O início de cada ciclo é a sístole atrial. Dura 0,1 segundos. No final da sístole atrial ocorre a diástole, assim como a sístole ventricular. A sístole ventricular dura 0,3 segundos. Durante a sístole, a pressão arterial aumenta nos ventrículos. Ao final da sístole ventricular, inicia-se a fase de relaxamento geral, com duração de 0,4 segundos. Em geral, o período de relaxamento dos átrios é de 0,7 segundos e o dos ventrículos é de 0,5 segundos. O significado fisiológico do período de relaxamento é que durante esse período ocorrem processos metabólicos entre as células e o sangue no miocárdio, ou seja, a capacidade de trabalho do músculo cardíaco é restaurada.
2 ... Características gerais dos órgãos respiratórios: cavidade nasal
A principal função da respiração é fornecer oxigênio aos tecidos humanos e liberá-los do dióxido de carbono. No sistema respiratório, distinguem-se os órgãos que realizam a condução aérea (cavidade nasal, nasofaringe, laringe, traquéia, brônquios) e funções respiratórias ou de troca gasosa (pulmões)
cavidade nasal
Distinguir entre o nariz externo e a cavidade nasal. Devido ao nariz externo, o volume da cavidade nasal aumenta. A cavidade nasal é dividida por um septo nasal vertical em duas metades simétricas, que na frente se comunicam com a atmosfera externa através do nariz externo com o auxílio de narina, e atrás - com a nasofaringe usando choan. Nas paredes laterais desta cavidade estão localizados cornetos, dividindo cada metade das passagens nasais da cavidade nasal. Abre na passagem nasal inferior ducto nasolacrimal através do qual uma certa quantidade de fluido lacrimal é liberada na cavidade nasal. As paredes da cavidade nasal são revestidas por uma membrana mucosa formada por epitélio ciliado.
A cavidade nasal é uma seção especializada do trato respiratório superior, pois aqui o ar inspirado é preparado para movimento adicional através do trato respiratório e fica exposto a tratamento especial:
aquece ou esfria até a temperatura corporal;
Hidratado por muco, localizado na mucosa nasal;
É limpo e desinfetado: o muco envolve as partículas de poeira que se depositam na membrana mucosa; o muco contém uma substância bactericida - lisozima, com a ajuda da qual as bactérias patogênicas são destruídas;
submetidos ao controle químico: na mucosa da parte superior da cavidade nasal estão localizados receptores olfativos.
A cavidade nasal possui cavidades adicionais seios paranasais localizado nos ossos aéreos do crânio: na mandíbula superior seio maxilar, no osso frontal - seio frontal (frontal), e também existem cavidades adicionais nos ossos esfenóide e etmóide. A inflamação da membrana mucosa desses seios leva a doenças graves sinusite e frontite.
Consideramos apenas um lado do processo respiratório - a respiração externa, ou seja, a troca de gases entre o corpo e o ambiente.
O local de consumo de oxigênio e de formação de dióxido de carbono são todas as células do corpo, onde ocorre a respiração tecidual ou interna. Por isso, quando se trata da respiração em geral, é necessário levar em consideração as formas e condições de transferência dos gases: oxigênio - dos pulmões para os tecidos, dióxido de carbono - dos tecidos para os pulmões. O intermediário entre as células e o meio ambiente é o sangue. Ele fornece oxigênio aos tecidos e remove o dióxido de carbono.
A movimentação dos gases do meio ambiente para o líquido e do líquido para o meio ambiente é realizada devido à diferença em sua pressão parcial. Um gás sempre se difunde de um ambiente onde há alta pressão, em um ambiente com menor pressão. Isso continua até que o equilíbrio dinâmico seja estabelecido.
Vamos traçar o caminho do oxigênio do meio ambiente para o ar alveolar, depois para os capilares da circulação pulmonar e sistêmica e para as células do corpo.
A pressão parcial de oxigênio no ar atmosférico é 21,1 kPa (158 mm Hg), no ar alveolar - 14,4-14,7 kPa (108-110 mm Hg) e no sangue venoso que flui para os pulmões, -5,33 kPa (40 mmHg ). No sangue arterial dos capilares da circulação sistêmica, a tensão de oxigênio é 13,6-13,9 kPa (102-104 mm Hg), no líquido intersticial - 5,33 kPa (40 mm Hg), nos tecidos - 2,67 kPa ( 20 mm Hg) e menos, dependendo da atividade funcional das células. Assim, em todas as etapas da movimentação do oxigênio, há uma diferença em sua pressão parcial, o que contribui para a difusão do gás.
O movimento do dióxido de carbono ocorre na direção oposta. A tensão do dióxido de carbono nos tecidos, nos locais de sua formação - 8,0 kPa ou mais (60 mm Hg ou mais), no sangue venoso - 6,13 kPa (46 mm Hg), no ar alveolar - 0,04 kPa (0,3mmHg). Portanto, a diferença na voltagem do dióxido de carbono ao longo de seu caminho é a causa da difusão do gás dos tecidos para o meio ambiente. O esquema de difusão dos gases através da parede dos alvéolos é mostrado na fig. 3. Contudo, o movimento dos gases não pode ser explicado apenas pelas leis físicas. Em um organismo vivo, nunca ocorre igualdade da pressão parcial de oxigênio e dióxido de carbono nas fases de seu movimento. Nos pulmões, há uma troca constante de gases devido aos movimentos respiratórios do tórax, enquanto nos tecidos a diferença na tensão dos gases é mantida por um processo contínuo de oxidação.
Arroz. 3. Esquema de difusão de gases através da membrana alveolar
Transporte de oxigênio no sangue. O oxigênio no sangue está em dois estados: dissolução física e ligação química com a hemoglobina. Dos 19% em volume de oxigênio extraído do sangue arterial, apenas 0,3% em volume está em estado dissolvido no plasma, enquanto o restante do oxigênio está quimicamente ligado à hemoglobina eritrocitária.
A hemoglobina forma com o oxigênio um composto muito frágil e de fácil dissociação - a oxiemoglobina: 1 g de hemoglobina liga-se a 1,34 ml de oxigênio. O conteúdo de hemoglobina no sangue é em média 140 g/l (14 g%). 100 ml de sangue podem ligar 14x1,34 = 18,76 ml de oxigênio (ou 19 vol%), que é basicamente a chamada capacidade de oxigênio do sangue. Portanto, a capacidade de oxigênio do sangue representa a quantidade máxima de oxigênio que pode ser ligada por 100 ml de sangue.
A saturação da hemoglobina com oxigênio varia de 96 a 98%. O grau de saturação da hemoglobina com oxigênio e a dissociação da oxiemoglobina (formação de hemoglobina reduzida) não são diretamente proporcionais à tensão de oxigênio. Esses dois processos não são lineares, mas seguem uma curva chamada curva de ligação ou dissociação da oxiemoglobina.
Na tensão zero de oxigênio, não há oxiemoglobina no sangue. Em valores baixos de pressão parcial de oxigênio, a taxa de formação de oxiemoglobina é baixa. A quantidade máxima de hemoglobina (45-80%) liga-se ao oxigênio em sua voltagem de 3,47-6,13 kPa (26-46 mm Hg). Um aumento adicional na tensão de oxigênio leva a uma diminuição na taxa de formação de oxiemoglobina.
A afinidade da hemoglobina pelo oxigênio diminui significativamente quando a reação do sangue muda para o lado ácido, que é observado nos tecidos e células do corpo devido à formação de dióxido de carbono. Esta propriedade da hemoglobina é essencial para o corpo. Nos capilares dos tecidos, onde a concentração de dióxido de carbono no sangue aumenta, a capacidade da hemoglobina de reter oxigênio diminui, o que facilita seu retorno às células. Nos alvéolos dos pulmões, onde parte do dióxido de carbono passa para o ar alveolar, a capacidade da hemoglobina de se ligar ao oxigênio aumenta novamente.
A transição da hemoglobina para a oxiemoglobina e desta para a reduzida também depende da temperatura. À mesma pressão parcial de oxigênio em ambiente a uma temperatura de 37-38 ° C, a maior quantidade de oxiemoglobina passa para a forma reduzida.
Assim, o transporte de oxigênio é fornecido principalmente devido à sua ligação química com a hemoglobina eritrocitária. A saturação da hemoglobina com oxigênio depende principalmente da pressão parcial do gás no ar atmosférico e alveolar. Uma das principais razões que contribuem para a liberação de oxigênio pela hemoglobina é a mudança da reação ativa do meio ambiente nos tecidos para o lado ácido.
Transporte de dióxido de carbono no sangue. A solubilidade do dióxido de carbono no sangue é maior que a solubilidade do oxigênio. No entanto, apenas 2,5-3% em volume de dióxido de carbono da sua quantidade total (55-58% em volume) está em estado dissolvido. A maior parte do dióxido de carbono está contida no sangue e nos eritrócitos na forma de sais de ácido carbônico (48-51% em volume), cerca de 4-5% em volume - em combinação com hemoglobina na forma de carbhemoglobina, cerca de 2/3 de todos os compostos de dióxido de carbono estão no plasma e cerca de 1/s nos eritrócitos.
O ácido carbônico é formado nos glóbulos vermelhos a partir do dióxido de carbono e da água. ELES. Sechenov foi o primeiro a sugerir que os eritrócitos deveriam conter algum fator, como um catalisador que acelerasse o processo de síntese do ácido carbônico. No entanto, somente em 1935 a suposição feita por I.M. Sechenov, foi confirmado. Foi agora estabelecido que os eritrócitos contêm anidrase carbônica (anidrase carbônica) - um catalisador biológico, uma enzima que acelera significativamente (300 vezes) a degradação do ácido carbônico nos capilares dos pulmões. Nos capilares dos tecidos, com a participação da anidrase carbônica, o ácido carbônico é sintetizado nos eritrócitos. A atividade da anidrase carbônica nos eritrócitos é tão grande que a síntese do ácido carbônico é acelerada dezenas de milhares de vezes.
O ácido carbônico remove bases da hemoglobina reduzida, resultando na formação de sais de ácido carbônico - bicarbonatos de sódio no plasma e bicarbonatos de potássio nos eritrócitos. Além disso, a hemoglobina forma um composto químico com o dióxido de carbono - a carbhemoglobina. Este composto foi descoberto pela primeira vez por I.M. Sechenov. O papel da carbhemoglobina no transporte de dióxido de carbono é bastante grande. Cerca de 25-30% do dióxido de carbono absorvido pelo sangue nos capilares da circulação sistêmica é transportado na forma de carbhemoglobina. Nos pulmões, a hemoglobina liga o oxigênio e passa para a oxiemoglobina. A hemoglobina reage com os bicarbonatos e desloca o ácido carbônico deles. O ácido carbônico livre é clivado pela anidrase carbônica em dióxido de carbono e água. O dióxido de carbono difunde-se através da membrana dos capilares pulmonares e passa para o ar alveolar. Uma diminuição na tensão do dióxido de carbono nos capilares dos pulmões contribui para a degradação da carbhemoglobina com liberação de dióxido de carbono.
Assim, o dióxido de carbono é transportado para os pulmões na forma de bicarbonatos e em estado de ligação química com a hemoglobina (carbemoglobina). Um papel importante nos mecanismos mais complexos de transporte de dióxido de carbono pertence à anidrase carbônica eritrocitária.
O objetivo final da respiração é fornecer oxigênio a todas as células e remover o dióxido de carbono do corpo. Para atingir esse objetivo de respiração, são necessárias uma série de condições:
1) atividade normal do aparelho respiratório externo e ventilação suficiente dos pulmões;
2) transporte normal de gases pelo sangue;
3) fornecer fluxo sanguíneo suficiente pelo sistema circulatório;
4) a capacidade dos tecidos de "captar" oxigênio do sangue que flui, utilizá-lo e liberar dióxido de carbono no sangue.
Assim, a respiração dos tecidos é proporcionada por relações funcionais entre os sistemas respiratório, sanguíneo e circulatório.
Índice da disciplina "Ventilação dos pulmões. Perfusão dos pulmões com sangue.":1. Ventilação dos pulmões. Ventilação pelo sangue dos pulmões. Espaço morto fisiológico. Ventilação alveolar.
2. Perfusão dos pulmões com sangue. Efeito da gravidade na ventilação dos pulmões. Efeito da gravidade na perfusão pulmonar com sangue.
3. Coeficiente de relação ventilação-perfusão nos pulmões. Troca gasosa nos pulmões.
4. Composição do ar alveolar. Composição gasosa do ar alveolar.
5. Tensão de gases nos capilares sanguíneos dos pulmões. A taxa de difusão de oxigênio e dióxido de carbono nos pulmões. Equação de Fick.
7. A afinidade da hemoglobina pelo oxigênio. Mudança na afinidade da hemoglobina pelo oxigênio. Efeito Bohr.
8. Dióxido de carbono. transporte de dióxido de carbono.
9. O papel dos eritrócitos no transporte de dióxido de carbono. Efeito Holden.
10. Regulação da respiração. Regulação da ventilação pulmonar.
A circulação sanguínea desempenha uma das funções mais importantes transferência de oxigênio dos pulmões para os tecidos e dióxido de carbono dos tecidos para os pulmões. O consumo de oxigênio pelas células dos tecidos pode variar significativamente, por exemplo, durante a transição do estado de repouso para a atividade física e vice-versa. Nesse sentido, o sangue deve ter grandes reservas necessárias para aumentar sua capacidade de transportar oxigênio dos pulmões para os tecidos e dióxido de carbono na direção oposta.
transporte de oxigênio.
A 37 C, a solubilidade de 02 em líquido é de 0,225 ml l-1 kPa-1 (0,03 ml/l/mm Hg). Sob condições de pressão parcial normal de oxigênio no ar alveolar, ou seja, 13,3 kPa ou 100 mm Hg, 1 l plasma sanguíneo pode transportar apenas 3 ml 02, o que não é suficiente para a vida do organismo como um todo. Em repouso, o corpo humano consome aproximadamente 250 ml de oxigênio por minuto. Para que os tecidos recebam tal quantidade de oxigênio em estado fisicamente dissolvido, o coração deve bombear uma grande quantidade de sangue por minuto. Na evolução dos seres vivos, o problema do transporte de oxigênio foi resolvido de forma mais eficaz devido à reversibilidade reação química com hemoglobina eritrocitária. O oxigênio é transportado pelo sangue dos pulmões para os tecidos do corpo pelas moléculas de hemoglobina, que estão contidas nos glóbulos vermelhos.
A hemoglobina é capaz de capturar oxigênio do ar alveolar (o composto é denominado oxiemoglobina) e libera a quantidade necessária de oxigênio nos tecidos. Uma característica da reação química do oxigênio com a hemoglobina é que a quantidade de oxigênio ligado é limitada pelo número de moléculas de hemoglobina nos glóbulos vermelhos. A molécula de hemoglobina possui 4 sítios de ligação de oxigênio que interagem de tal forma que a relação entre a pressão parcial de oxigênio e a quantidade de oxigênio transportada no sangue tem um formato de S, que é chamada de curva de saturação ou dissociação da oxihemoglobina (Fig. 10.18). A uma pressão parcial de oxigênio de 10 mm Hg. Arte. a saturação da hemoglobina com oxigênio é de aproximadamente 10% e em P02 30 mm Hg. Arte. - 50-60%. Com um novo aumento na pressão parcial de oxigênio de 40 mm Hg. Arte. até 60 mm Hg Arte. há uma diminuição na inclinação da curva de dissociação da oxiemoglobina e a porcentagem de sua saturação de oxigênio aumenta na faixa de 70-75 a 90%, respectivamente. Então a curva de dissociação da oxiemoglobina começa a assumir uma posição quase horizontal, à medida que aumenta a pressão parcial do oxigênio de 60 para 80 mm Hg. Arte. provoca um aumento na saturação da hemoglobina com oxigênio em 6%. Na faixa de 80 a 100 mm Hg. Arte. a porcentagem de formação de oxiemoglobina é de cerca de 2. Como resultado, a curva de dissociação da oxiemoglobina se transforma em uma linha horizontal e a porcentagem de saturação da hemoglobina com oxigênio atinge o limite, ou seja, 100. A saturação da hemoglobina com oxigênio sob a influência de P02 caracteriza uma espécie do "apetite" molecular deste composto por oxigênio.
Inclinação significativa da curva saturação da hemoglobina com oxigênio na faixa de pressão parcial de 20 a 40 mm Hg. Arte. contribui para o fato de que nos tecidos do corpo uma quantidade significativa de oxigênio pode se difundir do sangue nas condições de um gradiente de sua pressão parcial entre o sangue e as células do tecido (pelo menos 20 mm Hg). Uma porcentagem insignificante de saturação da hemoglobina com oxigênio na faixa de sua pressão parcial de 80 a 100 mm Hg. Arte. contribui para que uma pessoa sem risco de redução da saturação do sangue arterial com oxigênio possa se deslocar em altitudes de até 2.000 m acima do nível do mar.
 Arroz. 10.18. Curva de dissociação da oxihemoglobina. Limites de flutuação da curva em PC02 = 40 mm Hg. Arte. (sangue arterial) e PC02 = 46 mm Hg. Arte. (sangue venoso) mostram uma mudança na afinidade da hemoglobina pelo oxigênio ( Efeito Hoden).
Arroz. 10.18. Curva de dissociação da oxihemoglobina. Limites de flutuação da curva em PC02 = 40 mm Hg. Arte. (sangue arterial) e PC02 = 46 mm Hg. Arte. (sangue venoso) mostram uma mudança na afinidade da hemoglobina pelo oxigênio ( Efeito Hoden).
Reservas totais de oxigênio no corpo são devidos à sua quantidade, que está ligada aos íons Fe2 + na composição das moléculas orgânicas de hemoglobina nos eritrócitos e mioglobina nas células musculares.
Um grama de hemoglobina liga 1,34 ml 02. Portanto, normalmente, em uma concentração de hemoglobina de 150 g/l, cada 100 ml de sangue pode transportar 20,0 ml 02.
A quantidade de 02 que pode se ligar à hemoglobina dos glóbulos vermelhos quando saturada com 100% de sua quantidade é chamada capacidade de oxigênio da hemoglobina. Outro indicador da função respiratória do sangue é o conteúdo de 02 no sangue ( capacidade de oxigênio do sangue), o que reflete sua verdadeira quantidade, tanto associada à hemoglobina quanto fisicamente dissolvida no plasma. Como o sangue arterial normal está 97% saturado de oxigênio, 100 ml de sangue arterial contém aproximadamente 19,4 ml de O2.